Aby bolo možné komplexne hovoriť o hemoglobíne (Hb), je užitočné zaoberať sa najprv myoglobínom (Mb), ktorý je veľmi podobný hemoglobínu, ale je oveľa jednoduchší. Existujú prísne príbuzenské vzťahy medzi hemoglobínom a myoglobínom: obidva sú konjugované proteíny a ich protetická skupina (neproteínová časť) je skupina hemu .
Myoglobín je globulárny proteín pozostávajúci z jedného reťazca asi sto päťdesiatich aminokyselín (v závislosti od organizmu) a jeho molekulová hmotnosť je asi 18 Kd.
Ako je uvedené vyššie, je vybavená hemovou skupinou, ktorá je vložená do hydrofóbnej (alebo lipofilnej) časti proteínu, pozostávajúcej zo skladaní, ktoré možno pripísať štruktúram a-helixov vláknitých proteínov.
Myoglobín sa skladá prevažne zo segmentov α-helixov prítomných v počte ôsmich a pozostáva takmer výlučne z nepolárnych zvyškov (leucín, valín, metionín a fenylalanín), zatiaľ čo polárne zvyšky sú prakticky neprítomné (kyselina asparágová, kyselina glutámová, lyzín a arginín); Jedinými polárnymi zvyškami sú dva histidíny, ktoré zohrávajú zásadnú úlohu pri napadnutí kyslíka do hemovej skupiny.
Hemová skupina je chromoforová skupina (absorbuje sa vo viditeľnom) a je funkčnou skupinou myoglobínu.
Pozri tiež: glykovaný hemoglobín - hemoglobín v moči
Trochu chémie
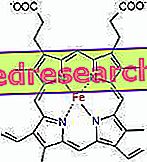
Spojenie medzi protoporfyrínom a železom je typická väzba takzvaných koordinačných zlúčenín, ktoré sú chemickými zlúčeninami, v ktorých centrálny atóm (alebo ión) tvorí väzby s inými chemickými druhmi v množstve vyššom ako je jeho oxidačné číslo (elektrický náboj). V prípade hemu sú tieto väzby reverzibilné a slabé.
Koordinačné číslo (počet koordinačných väzieb) železa je šesť: okolo železa môže byť šesť molekúl, ktoré zdieľajú väzbové elektróny.
Ak chcete vytvoriť koordinačnú zlúčeninu, trvá dva orbitály so správnou orientáciou: jeden je schopný "kúpiť" elektróny a druhý je schopný darovať.
V heme tvorí železo štyri rovinné väzby so štyrmi atómami dusíka, ktoré sú v strede proto-porfyrínového kruhu a piata väzba s proximálnym histidínovým dusíkom; železo má šieste voľné koordinačné spojenie a môže sa viazať na kyslík.
Keď je železo vo forme voľného iónu, jeho d- orbitály majú rovnakú energiu; v myoglobíne sa ión železa viaže na protoporfyrín a histidín: tieto druhy magneticky rušia orbitály železa; rozsah odchýlky sa bude líšiť pre rôzne orbitály podľa ich priestorovej orientácie a rušivých druhov. Keďže celková energia orbitálov sa musí vrátiť na konštantnú hodnotu, rušenie spôsobuje energetickú separáciu medzi rôznymi orbitálmi: energia získaná niektorými orbitálmi je ekvivalentná energii stratenej ostatnými.
Ak oddelenie, ktoré nastane medzi orbitálmi, nie je veľmi veľké, uprednostňuje sa vysoko spinové elektronické usporiadanie: väzbové elektróny sa snažia usporiadať v paralelných roztočeniach v niekoľkých možných podúrovniach (maximálna multiplicita); ak je na druhej strane porucha veľmi silná a existuje veľké oddelenie medzi orbitálmi, môže byť vhodnejšie spárovať väzbové elektróny v nižších energetických orbitáloch (nízke odstreďovanie).
Keď sa železo viaže na kyslík, molekula má nízku spinovú dispozíciu, zatiaľ čo keď má železo šiestu voľnú koordinačnú väzbu, molekula má vysokú rotáciu.
Vďaka tomuto rozdielu v rotácii, prostredníctvom spektrálnej analýzy myoglobínu, sme schopní pochopiť, či je viazaný kyslík (MbO2) alebo nie (Mb).
Myoglobín je proteín typický pre svaly (ale nie len vo svaloch).
Myoglobín sa získava zo spermií, v ktorých je prítomný vo veľkých množstvách a potom sa čistí.
Cetacea majú dýchanie podobné ako ľudské bytosti: majú pľúca, ktoré musia absorbovať vzduch dýchacím procesom; veľryba spermie musí niesť čo najviac kyslíka vo svaloch, ktoré sú schopné hromadiť kyslík tým, že ho viaže na prítomný myoglobín; kyslík sa potom pomaly uvoľňuje, keď je kytovec ponorený, pretože jeho metabolizmus vyžaduje kyslík: čím väčšie je množstvo kyslíka, ktoré je sperma schopná absorbovať a tým viac kyslíka je k dispozícii počas ponoru.

Aby bolo mäso viac či menej červené, je obsah hemoproteínov (je to hem, ktorý robí mäso červené).
Hemoglobín má mnoho štruktúrnych analógií s myoglobínom a je schopný viazať molekulárny kyslík reverzibilne; ale zatiaľ čo myoglobín sa vo všeobecnosti obmedzuje na svaly a periférne tkanivá, hemoglobín sa nachádza v erytrocytoch alebo červených krvinkách (sú to pseudo bunky, to znamená, že nie sú reálnymi bunkami), ktoré tvoria 40% krvi.
Hemoglobín je tetrameter, to znamená, že sa skladá zo štyroch polypeptidových reťazcov, z ktorých každý má hémovú skupinu a dva po dvoch (v ľudskej bytosti sú dva alfa reťazce a dva beta reťazce).
Hlavnou funkciou hemoglobínu je transport kyslíka ; ďalšou funkciou krvi, v ktorej sa hemoglobín zúčastňuje, je transport látok do tkanív.
V ceste z pľúc (bohatej na kyslík) do tkanív, hemoglobín nesie kyslík (zároveň iné látky tiež zasahujú do tkanív), zatiaľ čo v opačnom smere so sebou nesie odpad zozbieraný z tkanív, najmä oxidu uhličitého produkovaného v metabolizme.
Pri vývoji ľudskej bytosti existujú gény, ktoré sú exprimované len po určitú dobu; preto máme rôzne hemoglobíny: fetálny, embryonálny, dospelého muža.
Reťaze, ktoré tvoria tieto rôzne hemoglobíny majú rôzne štruktúry, ale s podobnosťou funkcie, ktorú vykonávajú, sú viac-menej rovnaké.
Vysvetlenie prítomnosti niekoľkých rôznych reťazcov je nasledovné: počas evolučného procesu organizmov sa tiež vyvinul hemoglobín, ktorý sa špecializuje na prepravu kyslíka z oblastí bohatých na deficitné oblasti. Na začiatku evolučného reťazca hemoglobín prenášal kyslík v malých organizmoch; v priebehu evolúcie organizmy dosiahli väčšie rozmery, preto sa hemoglobín zmenil tak, aby bol schopný transportovať kyslík do oblastí vzdialenejších od miesta, kde bol bohatý; na to sa v priebehu evolučného procesu kodifikovali nové štruktúry reťazcov tvoriacich hemoglobín.
Myoglobín viaže kyslík aj pri miernom tlaku; v periférnych tkanivách je tlak (PO2) asi 30 mmHg: pri tomto tlaku myoglobín neuvoľňuje kyslík, takže by bol neúčinný ako nosič kyslíka. Na druhej strane má hemoglobín pružnejšie správanie: viaže kyslík pri vysokých tlakoch a uvoľňuje ho, keď tlak klesá.
Keď je proteín funkčne aktívny, môže mierne zmeniť svoj tvar; napríklad okysličený myoglobín má inú formu ako neokyslený myoglobín a táto mutácia neovplyvňuje susedné.
Diskurz je odlišný v prípade asociovaných proteínov, ako je hemoglobín: keď je reťazec okysličený, je indukovaný na zmenu jeho tvaru, ale táto modifikácia je trojrozmerná, preto sú ovplyvnené aj iné reťazce tetrametra. Skutočnosť, že reťazce sú navzájom spojené, vedie k domnienke, že zmena jedného ovplyvňuje aj tie susedné, aj keď v inom rozsahu; keď je reťazec okysličený, ostatné reťazce tetrametra preberajú „menej nepriateľský postoj“ voči kyslíku: ťažkosti, s ktorými sa reťazce okysličovadiel znižujú, keď sa reťazce nachádzajú vedľa seba. Rovnaký argument platí pre deoxygenáciu.
Kvartérna štruktúra deoxyhemoglobínu sa nazýva T (napätá) forma, zatiaľ čo oxyhemoglobín sa nazýva R forma (uvoľnená); v natiahnutom stave existuje rad pomerne silných elektrostatických interakcií medzi kyslými aminokyselinami a bázickými aminokyselinami, ktoré vedú k rigidnej štruktúre deoxyhemoglobínu (to je dôvod, prečo "napätá forma"), zatiaľ čo keď je viazaný kyslík, entita týchto interakcie klesajú (to je dôvod, prečo "uvoľnená forma"). Okrem toho v neprítomnosti kyslíka je náboj histidínu (pozri štruktúru) stabilizovaný opačným nábojom kyseliny asparágovej, zatiaľ čo v prítomnosti kyslíka existuje tendencia proteínu strácať protón; to všetko znamená, že okysličený hemoglobín je silnejšia kyselina ako deoxygenovaná hemoglobia: bohr efekt .
V závislosti od pH sa skupina hemu viaže na kyslík viac alebo menej: v kyslom prostredí hemoglobín uvoľňuje kyslík ľahšie (napätá forma je stabilná), zatiaľ čo v základnom prostredí je väzba s kyslíkom silnejší.

Každý hemoglobín uvoľní 0, 7 protónov na prichádzajúci mol kyslíka (O2).
Bohrov efekt umožňuje hemoglobínu zlepšiť jeho schopnosť prenášať kyslík.
Hemoglobín, ktorý robí cestu z pľúc do tkanív, sa musí vyrovnať podľa tlaku, pH a teploty.
Vidíme vplyv teploty .
Teplota v pľúcnych alveolách je asi o 1-1, 5 ° C nižšia ako vonkajšia teplota, zatiaľ čo vo svaloch je teplota asi 36, 5-37 ° C; S rastúcou teplotou sa saturačný faktor znižuje (s rovnakým tlakom): toto sa deje preto, že sa zvyšuje kinetická energia a disociácia.

Existujú aj iné faktory, ktoré môžu ovplyvniť schopnosť hemoglobínu viazať sa na kyslík, jedným z nich je koncentrácia 2, 3-bisfosfoglycerátu .
2, 3 bisfosfoglycerát je metabolické činidlo prítomné v erytrocytoch v koncentrácii 4 až 5 mM (v žiadnej inej časti tela nie je prítomná v takej vysokej koncentrácii).
Pri fyziologickom pH je 2, 3 bisfosfoglycerát deprotonovaný a má na ňom päť negatívnych nábojov; to ide do klinu medzi dvoma beta reťazcami hemoglobínu, pretože tieto reťazce majú vysokú koncentráciu pozitívnych nábojov. Elektrostatické interakcie medzi beta reťazcami a bisfosfoglycerátovými reťazcami dodávajú systému určitú tuhosť: získava sa napnutá štruktúra, ktorá má malú afinitu k kyslíku; počas okysličovania sa potom vylučuje 2, 3 bisfosfoglycerát.
V erytrocytoch sa nachádza špeciálny prístroj, ktorý premieňa 1, 3-bisfosfoglycerát (produkovaný metabolizmom) na 2, 3-bisfosfoglycerát tak, aby dosiahol koncentráciu 4-5mM, a preto je hemoglobín schopný výmeny l. kyslíka v tkanivách.
Hemoglobín prichádzajúci do tkaniva je v uvoľnenom stave (viazaný na kyslík), ale v blízkosti tkaniva, je karboxylovaný a prechádza do napnutého stavu: proteín v tomto stave má nižšiu tendenciu viazať sa kyslíkom, v porovnaní s uvoľneným stavom, preto hemoglobín uvoľňuje kyslík do tkaniva; okrem toho, v dôsledku reakcie medzi vodou a oxidom uhličitým, dochádza k produkcii iónov H +, a tým k ďalšiemu kyslíku v dôsledku bohr efektu.
Oxid uhličitý difunduje do erytrocytu prechádzajúceho cez plazmatickú membránu; vzhľadom na to, že erytrocyty tvoria približne 40% krvi, mali by sme očakávať, že do nich vstúpi len 40% oxidu uhličitého, ktorý difunduje z tkanív, v skutočnosti 90% oxidu uhličitého vstupuje do červených krviniek, pretože obsahujú enzým, ktorý konvertuje oxidu uhličitého v kyseline uhličitej vyplýva, že stacionárna koncentrácia oxidu uhličitého v erytrocytoch je nízka, a preto je rýchlosť vstupu vysoká.

Ďalší fenomén, ktorý sa vyskytuje, keď erytrocyt dosiahne tkanivo, je nasledovný: gradientom, HCO3- (derivát oxidu uhličitého) vychádza z erytrocytu a na vyváženie výstupu záporného náboja máme vstup chloridov, ktoré určujú zvýšenie osmotického tlaku: na vyrovnanie tejto odchýlky je tiež vstup vody, ktorý spôsobuje opuch erytrocytov (účinok HAMBURGER). Opačný jav nastáva, keď erytrocyt dosiahne pľúcne alveoly: dochádza k deflácii erytrocytov (účinok HALDANE). Venózne erytrocyty (priamo do pľúc) sú teda okrúhle ako arteriálne.


