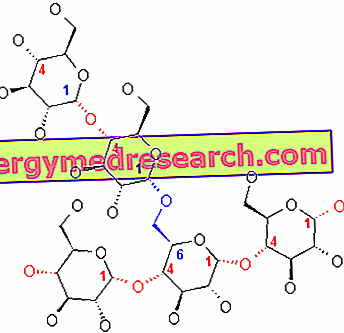
Glykogén je makromolekula (molekulová hmotnosť asi 400 miliónov daltonov) a-glukózy, v ktorej sú prevažne a-1, 4 glykozidové väzby a vetvy v pomere 1:10, vďaka a-1, 6 glykozidovým väzbám.
Glykogén predstavuje rezervný materiál a je kontinuálne degradovaný a rekonštituovaný; vo všetkých telesných bunkových hmotách, máme asi 100 g glykogénu: väčšina z nich je v pečeni, kde je mobilná, a preto môže byť použitá ako rezerva pre ostatné orgány (glykogén vo svaloch nie je mobilný).
Enzýmy, ktoré katalyzujú degradáciu a syntézu glykogénu, sú všetky v cytoplazme, preto je potrebný regulačný systém, ktorý robí jednu cestu neaktívnou, keď je druhá aktívna: ak je k dispozícii glukóza, táto je premenená na glykogén (anabolizmus), ktorý je rezervou, naopak, ak sa vyžaduje glukóza, potom sa glykogén degraduje (katabolizmus).
Enzýmom, ktorý sa podieľa najmä na degradácii glykogénu, je glykogénfosforyláza ; tento enzým je schopný štiepiť a-1, 4 glykozidovú väzbu použitím, ako lytického činidla, anorganického ortofosforečnanu: získa sa štiepenie prostredníctvom fosforolytického a glukózového 1-fosfátu.
Pri piatich alebo šiestich jednotkách z miesta rozvetvenia enzým glykogénfosforyláza už nie je schopný pôsobiť, preto sa oddelí od glykogénu a je nahradený enzýmom, ktorý je transferázou : v katalytickom mieste tohto enzýmu je 'histidín, ktorý umožňuje prenos troch sacharidových jednotiek na najbližší glykozidový reťazec (histidín napáda prvý uhlík molekuly glukózy). Práve zmieneným enzýmom je glykozyltransferáza ; na konci pôsobenia tohto enzýmu zostáva na bočnom reťazci iba jedna glukózová jednotka s prvým uhlíkom naviazaným na šiesty uhlík glukózy v hlavnom reťazci. Posledná glukózová jednotka bočného reťazca sa uvoľňuje pôsobením enzýmu a-1, 6 glykozidázy (tento enzým je druhou časťou enzýmu s odštiepením); vzhľadom na to, že vetvy v glykogéne sú v pomere 1:10, od úplnej degradácie makromolekuly získame približne 90% glukóza-1-fosfátu a približne 10% glukózy.
Pôsobenie vyššie uvedených enzýmov umožňuje elimináciu bočného reťazca z molekuly glykogénu; aktivita týchto enzýmov sa môže opakovať, až kým nedôjde k úplnej degradácii reťazca.
Uvažujme o hepatocyte; glukóza (asimilovaná cez diétu), keď vstupuje do bunky, je premenená na glukózo-6-fosfát a je teda aktivovaná. 6-fosfát glukózy, pôsobením fosfoglukomutázy, sa transformuje na glukóza-1-fosfát: posledne uvedený nie je bezprostredným prekurzorom biosyntézy; v biosyntéze sa používa aktivovaná forma cukrov, ktorá je reprezentovaná cukrom viazaným na difosfát: zvyčajne uridyldiposfát (UDP). Glukóza 1-fosfát sa potom premení na UDP-glukózu; tento metabolit je pod vplyvom glykogénsyntázy, ktorá je schopná viazať UDP-glukózu na neredukujúci koniec rastúceho glykogénu: predĺžený glykogén sa získava z glukozidovej jednotky a UDP. UDP je konvertovaný enzýmom nukleosidedifosphokinase na UTP, ktorý sa vracia do obehu.
Degradácia glykogénu nastáva pôsobením glykogénfosforylázy, ktorá uvoľňuje molekulu glukózy a transformuje ju na glukózový 1-fosfát. Následne fosfoglukomutáza konvertuje glukózový 1-fosfát na glukózový 6-fosfát.
Glykogén sa syntetizuje predovšetkým v pečeni a vo svaloch: v tele sa nachádza 1-1, 2 g glykogénu distribuovaného v svalovej hmote.
Glykogén myocytu predstavuje rezervu energie len pre túto bunku, zatiaľ čo glykogén obsiahnutý v pečeni je tiež rezervou pre ostatné tkanivá, to znamená, že môže byť poslaný ako glukóza do iných buniek.
Glukóza-6-fosfát získaný vo svaloch z degradácie glykogénu sa potom v prípade potreby energie posiela na glykolýzu; v pečeni sa glukóza-6-fosfát premieňa na glukózu pôsobením glukóza-6-fosfátovej fosfatázy (enzýmová charakteristika hepatocytov) a dopravuje sa do obežného prúdu.
Ako glykogénsyntáza, tak glykogénfosforyláza fungujú na neredukujúcich jednotkách glykogénu, takže musí existovať hormonálny signál, ktorý riadi aktiváciu jednej cesty a blokovanie druhého (alebo naopak).
V laboratóriu bolo možné predĺžiť glykogénový reťazec, využiť glykogénfosforylázu a používať glukóza-1-fosfát vo veľmi vysokej koncentrácii.
V bunkách glykogénfosforyláza katalyzuje len degradačnú reakciu, pretože koncentrácie metabolitov sú také, že posunú rovnováhu nasledujúcej reakcie doprava (tj smerom k degradácii glykogénu):

Pozrime sa na mechanizmus pôsobenia glykogénfosforylázy: kyslíkový kyslík (ktorý pôsobí ako most medzi jednotkami glukózy) sa viaže na fosforylový vodík: reakčný medziprodukt je tvorený karbokationom (na glukóze, ktorá je končatiny), na ktoré sa fosforyl (Pi) viaže veľmi rýchlo.
Glykogénfosforyláza vyžaduje kofaktor, ktorým je pyridoxal fosfát (táto molekula je tiež kofaktorom pre transaminázy): má fosforyl protónovaný iba čiastočne (pyridoxal fosfát je obklopený hydrofóbnym prostredím, ktoré odôvodňuje prítomnosť protónov viazaných na neho)., Fosforyl (Pi) je schopný poskytnúť protón glykogénu, pretože takéto fosforyly sa potom získajú, protón z čiastočne protónovaného fosforylu pyridoxal fosfátu. Pravdepodobnosť, že pri fyziologickom pH fosfory stráca protón a zostáva úplne deprotonovaná, je veľmi nízka.
Teraz sa pozrime, ako funguje fosfoglukomutáza. Tento enzým má fosforylovaný serínový zvyšok v katalytickom mieste; serín poskytuje fosforylu na glukózový 1-fosfát (v polohe šesť): glukóza 1, 6-bisfosfát sa vytvára na krátku dobu, potom sa serín fosforyluje tým, že sa fosforyl v polohe jedna. Fosfoglukus mutáza môže pôsobiť v oboch smeroch, t.j. premieňať glukózový 1-fosfát na glukózový 6-fosfát alebo naopak; ak sa vyrába glukóza-6-fosfát, môže sa priamo poslať na glykolýzu, do svalov alebo transformovať na glukózu v pečeni.
Enzým uridylfosfoglukus transferáza (alebo UDP glukóza pyrofosforyláza) katalyzuje prenosovú reakciu glukózového 1-fosfátu na UTP fosforylovým atakom a.
Práve opísaný enzým je pyrofosforyláza: tento názov je spôsobený skutočnosťou, že opačná reakcia, ktorá bola práve opísaná, je pyrofosforylácia.
UDP glukóza, získaná ako je opísané, je schopná predĺžiť glykogénový reťazec monosacharidovej jednotky.
Reakciu je možné vyvíjať smerom k tvorbe UDP glukózy odstránením produktu, ktorým je pyrofosfát; enzým pyrofosfatáza konvertuje pyrofosfát na dve molekuly ortofosforečnanu (hydrolýza anhydridu) a tým udržuje koncentráciu pyrofosforečnanu tak nízku, že proces tvorby glukózy UDP je uprednostňovaný termodynamicky.
Ako bolo uvedené, glukóza UDP je vďaka pôsobeniu glykogénsyntázy schopná predĺžiť glykogénový reťazec.
Dôsledky (v pomere 1:10) sú spôsobené tým, že keď glykogénový reťazec pozostáva z 20-25 jednotiek, zasahuje vetviaci enzým (majúci histidín na svojom katalytickom mieste), schopný prenosu série 7 -8 glykozidických jednotiek ďalej po 5-6 jednotkách: vzniká tak nové vetvenie.
Z dôvodov nervového pôvodu alebo ak je energia potrebná kvôli fyzickej námahe, adrenalín sa vylučuje z nadobličiek.
Cieľovými bunkami adrenalínu (a norepinefrínu) sú bunky pečene, svalov a tukového tkaniva (v druhom prípade dochádza k degradácii triglyceridov a cirkulácii mastných kyselín: v mitochondriách je teda produkovaná glukóza 6). -fosfát, ktorý sa má poslať na glykolýzu, zatiaľ čo v adipocytoch sa glukózový 6-fosfát transformuje na glukózu pôsobením enzýmu glukóza-6-fosfát fosfatázy a vyváža do tkanív).
Pozrime sa teraz na spôsob pôsobenia adrenalínu. Adrenalín sa viaže na receptor umiestnený na bunkovej membráne (myocytov a hepatocytov), čo určuje transláciu signálu zvonku dovnútra bunky. Aktivuje sa proteínkináza, ktorá pôsobí súčasne na systémy, ktoré regulujú syntézu a degradáciu glykogénu:
Glykogénsyntáza existuje v dvoch formách: defosforylovaná forma (aktívna) a fosforylovaná forma (inaktívna); proteínkináza fosforyluje glykogénsyntázu a blokuje jej pôsobenie.
Glykogénfosforyláza môže existovať v dvoch formách: aktívna forma, v ktorej je prítomný fosforylovaný serín a inaktívna forma, v ktorej je serín defosforylovaný. Glykogénfosforyláza môže byť aktivovaná enzýmom glykogénfosforylázkinázy . Glykogénfosforylázkináza je aktívna, ak je fosforylovaná a neaktívna, ak je defosforylovaná; proteínkináza má ako substrát kinázu glykogénfosforylázy, t.j. je schopná fosforylovať (a teda aktivovať) posledne menovanú, ktorá zase aktivuje glykogénfosforylázu.
Akonáhle je adrenalínový signál u konca, účinok, ktorý má v bunke, musí tiež skončiť: fosfatázové enzýmy zasiahnu, potom na proteínové druhy.